Г.Ю.Ризниченко
Лекции по математическим моделям в биологии
ЛЕКЦИЯ 8
КОЛЕБАНИЯ
В БИОЛОГИЧЕСКИХ СИСТЕМАХ
Понятие автоколебаний. Изображение автоколебательной системы на фазовой плоскости. Предельные циклы. Условия существования предельных циклов. Рождение предельного цикла. Бифуркация Андронова ‑ Хопфа. Мягкое и жесткое возбуждение колебаний. Модель брюсселятор. Примеры автоколебательных моделей процессов в живых системах. Колебания в темновых процессах фотосинтеза. Автоколебания в модели гликолиза. Внутриклеточные колебания концентрации кальция. Клеточные циклы.
Для биологических систем характерно периодическое изменение различных характеристик. Период этих колебаний может быть связан с периодическими изменениями условий жизни на Земле - смена времен года, смена дня и ночи. Существуют и другие геофизические ритмы –солнечные, лунные, связанные с периодами атмосферных явлений. Геофизические и биологические ритмы сопоставлены на рис. 8.1. Но многие периодические процессы имеют частоту изменения, не связанную очевидным образом с внешними геокосмическими циклами. Это так называемые «биологические часы» различной природы, начиная от колебаний биомакромолекул, биохимических колебаний, вплоть до популяционных волн.
Внутриклеточные колебания
задают эндогенные биологические ритмы, которые свойственны всем живым системам.
Именно они определяют периодичность деления клеток, отмеряют время рождения и
смерти живых организмов. Модели колебательных систем используются в ферментативном
катализе, теории иммунитета, в теории трансмембранного ионного переноса,
микробиологии и биотехнологии.
С некоторыми из типов периодических движений мы уже имели дело при
рассмотрении особых точек типа центр и затухающих или нарастающих колебаний в
случае устойчивого и неустойчивого фокуса. Однако «биологические часы» имеют
свойство, отличающее их от рассмотренных типов колебаний - неизменность во времени периода и амплитуды
таких колебаний, означающую стационарность
и устойчивость колебательного режима.

Рис. 8.1.
Космофизические, геофизические и биологические ритмы. Справа – шкала
периодов, слева – шкала частот
В
данном случае периодическое изменение величин представляет собой один из типов
стационарного поведения системы. Если колебания в системе имеют постоянные
период и амплитуду, устанавливаются независимо от начальных условий и
поддерживаются благодаря свойствам самой системы, а не вследствие воздействия
периодической силы, система называется автоколебательной.
Незатухающие колебания в таких системах устойчивы, так как отклонения от стационарного колебательного режима затухают. К классу автоколебательных систем относятся колебания в гликолизе и других метаболических системах, периодические процессы фотосинтеза, колебания концентрации кальция в клетке, колебания численности животных в популяциях и сообществах.
Предельный
цикл. В фазовом пространстве такому типу поведения
соответствует притягивающее множество (аттрактор), называемое предельным циклом.
Предельный цикл есть
изолированная замкнутая кривая на фазовой плоскости, к которой в пределе при t ® ¥ стремятся все интегральные кривые. Предельный цикл
представляет стационарный режим с определенной амплитудой, не зависящий от
начальных условий, а определяющийся только организацией системы. Существование
предельного цикла на фазовой плоскости есть основной признак автоколебательной
системы. Очевидно, что при автоколебательном процессе фаза колебаний может быть
любой.
Остановимся на общих характеристиках автоколебательных систем. Рассмотрим систему уравнений общего вида:
![]()
![]()
Если T
(T > 0) — наименьшее число, для которого при всяком t
![]()
![]()
то
изменение переменных x = x(t), y = y(t) называется
периодическим изменением с периодом T.
Периодическому изменению соответствует замкнутая траектория на фазовой плоскости, и обратно: всякой замкнутой траектории соответствует бесконечное множество периодических изменений, отличающихся друг от друга выбором начала отсчета времени.
Если периодическому изменению на фазовой плоскости соответствует изолированная замкнутая кривая, к
которой с внешней и внутренней стороны приближаются (при возрастании t) соседние траектории по спиралям, эта изолированная замкнутая траектория есть
предельный цикл.
Простые примеры позволяют убедиться, что система общего вида (8.1) допускает в качестве траекторий предельные циклы.
Например, для системы
![]()
![]()
траектория
![]()
![]()
![]()
а уравнения всех других фазовых траекторий запишутся в виде:
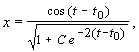

Значениям постоянной интегрирования С > 0 соответствуют фазовые траектории, накручивающиеся на предельный
цикл изнутри (при t ® ¥), а значениям –1 < C < 0 траектории, накручивающиеся
снаружи.
Предельный цикл называется устойчивым, если существует такая область
на фазовой плоскости, содержащая этот предельный цикл, - окрестность e, что все фазовые траектории, начинающиеся в окрестности
e, асимптотически при t ® ¥ приближаются к предельному циклу.
Если же, наоборот, в любой сколь угодно малой окрестности e
предельного цикла существует по крайней мере одна фазовая траектория, не
приближающаяся к предельному циклу при
t ® ¥, то
такой предельный цикл называется неустойчивым. Такие циклы разделяют области влияния (бассейны) разных
притягивающих множеств.
На рис. 8.2 изображены устойчивый предельный цикл (а) и неустойчивые (б) и (в). Рис. 8.2. Устойчивый (а)
и неустойчивые (б и в) предельные циклы на фазовой
плоскости
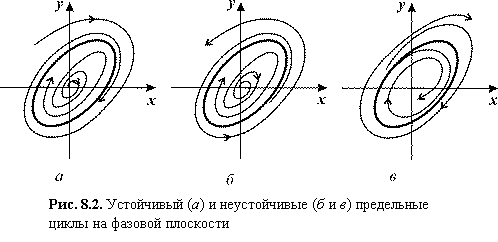
Неустойчивые предельные циклы, подобные
изображенному на рис. 8.2 б, такие, что все траектории с одной стороны (например, изнутри)
приближаются к ним, а с другой стороны (например, извне) удаляются от них при t ® ¥, называют
«полуустойчивыми» или двойными. Последнее название связано с тем, что обычно
такие циклы при подходящем изменении параметра системы расщепляются на два,
один из которых устойчив, а другой неустойчив.
А.М. Ляпунов показал, что для исследования устойчивости периодического движения x = j(t), y = y(t) можно идти по пути линеаризации уравнений, подобно тому, как мы это делали при исследовании устойчивости состояний равновесия. Если положить
![]()
![]()
подставить эти выражения в уравнения (8.1), разложить правые части этих уравнений - функции
![]()
![]()
в ряды
по степеням x и h и
отбросить нелинейные члены, то мы получим линейные уравнения (уравнения первого
приближения) для координат возмущения x и h:
![]()
![]()
Коэффициенты в правой части:
![]()
![]()
![]()
![]()
Это система линейных дифференциальных уравнений с
периодическими коэффициентами периода T,
поскольку a, b, c, d суть функции от j, y — периодических
функций времени с периодом T. Общий
вид ее решения
![]()
![]()
Здесь ![]()
![]()
![]()

где x = j(t), y = y(t) — любое периодическое решение,
соответствующее рассматриваемому предельному циклу, T — период решения.
Таким образом, устойчивость предельного цикла (и устойчивость в смысле
Ляпунова соответствующих периодических движений) определяется знаком
характеристического показателя. Предельный цикл устойчив, если h < 0 и неустойчив,
если h > 0. Если
же h = 0,
уравнения первого приближения не решают вопроса об устойчивости периодического
движения.
Для нахождения предельных циклов не существует таких простых аналитических методов, как для нахождения стационарных точек и исследования их устойчивости. Однако, исследование фазовой плоскости системы позволяет ответить на вопрос, есть в данной системе предельный цикл, или нет.
Сформулируем несколько теорем, определяющих наличие предельного цикла по
топологическому строению фазовой плоскости. Они могут быть полезны как при
аналитическом, так и при компьютерном анализе системы.
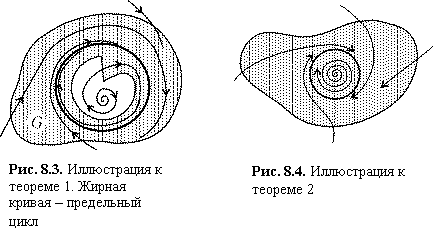
Теорема 1. Пусть на фазовой
плоскости существует область, из которой фазовые траектории не выходят, и в
которой нет положений равновесия (особых точек). Тогда в этой области
обязательно существует предельный цикл, причем все остальные траектории
обязательно наматываются на него.
На рис. 8.3. изображена такая область G, из которой фазовые траектории не выходят. Это означает, что фазовые траектории либо входят, пересекая границу, внутрь области, либо сама граница является траекторией. Легко видеть, что такая область не может быть односвязной. Поскольку траектория наматывается на предельный цикл изнутри, это означает, что внутри этого предельного цикла на фазовой плоскости существует либо неустойчивая особая точка, либо неустойчивый предельный цикл, очевидно, не принадлежащие рассматриваемой области G.
Таким образом, если найти на фазовой плоскости такую двусвязную область, что направления фазовых траекторий на всей границе обращены внутрь этой области, то можно утверждать, что внутри этой области имеется предельный цикл.
Теорема 2. Если
существует на фазовой плоскости некоторая замкнутая область, такая, что все
фазовые траектории, пересекающие границу этой области, входят в нее, и внутри
этой области находится неустойчивая особая точка, то в этой области обязательно
имеется хотя бы один предельный цикл (рис. 8.4)
Рис. 8.4. Иллюстрация к теореме 2 Рис. 8.3. Иллюстрация к теореме
1. Жирная кривая – предельный цикл
Приведем
также некоторые критерии отсутствия замкнутых фазовых траекторий (в том числе
предельных циклов).
1. Если в системе не существует особых точек, то в ней не может быть и
замкнутых фазовых траекторий.
2. Если в системе существует только одна особая точка, отличная от узла,
фокуса и центра (например, седло), то такая система не допускает замкнутых
фазовых траекторий.
3. Если в системе имеются только простые особые точки, причем через все точки типа узел и фокус проходят интегральные кривые, уходящие на бесконечность, то в такой системе нет замкнутых фазовых траекторий.
В случае, если критерии 1–3 выполнены, можно с уверенностью утверждать, что в системе нет предельных циклов. Однако невыполнение этих критериев еще не позволяет сделать вывод о наличии в системе предельных циклов и, следовательно, автоколебаний.
Рис. 8.5. Фазовый портрет системы, имеющий
устойчивый и неустойчивый (пунктир) предельные циклы

Неустойчивый предельный цикл также может содержаться в фазовом портрете грубых
систем. Однако такой предельный цикл не соответствует реальному периодическому
процессу, он играет лишь роль «водораздела», по обе стороны которого траектории
имеют различное поведение. Например, на рис. 8.5 представляет собой
сепаратрису, отделяющую область тяготения траекторий к устойчивой особой точке,
с одной стороны, и к устойчивому предельному циклу, с другой.
Рождение предельного цикла. Бифуркация Андронова-Хопфа.
Существование предельных циклов возможно лишь в системе типа (8.1), правые части которой представлены нелинейными функциями.
На бифуркационной диаграмме 4.11 мы видели, что при пересечении оси абсцисс происходит смена устойчивости фокуса. Нулевым значениям действительной части характеристических чисел (ляпуновских показателей) соответствует особая точка типа центр. В нелинейной системе, где возникает неустойчивый фокус, при этом возможно рождение предельного цикла. Такой переход легко проследить в «модельной» системе:
![]()
![]()
Схематически возникновение предельного цикла в системе (8.3) изображено на фазопараметрической диаграмме на рис. 8.6.
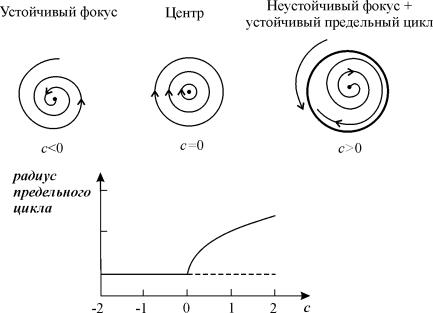
Рис. 8.6. Закритическая (суперкритическая) бифуркация
Андронова-Хопфа. Мягкое возбуждение. При с>0 возникают автоколебания,
амплитуда которых растет с увеличением с.
Выполнению условия Re l1,2 = 0, причем Im l1,2 ¹ 0, соответствует бифуркация Андронова–Хопфа или бифуркация рождения (исчезновения) предельного цикла. Бифуркация впервые была исследована А.А. Андроновым для случая N = 2 и обобщена Е. Хопфом на системы с произвольной размерностью. (Андронов А.А., Витт А.А., Хайкин С.Э. Теория колебаний. М., Наука, 1981; Hopf E., 1942)
Существуют два типа бифуркации Андронова–Хопфа. Только что мы рассмотрели суперкритическую бифуркацию (мягкое возбуждение автоколебаний). Возможна также субкритическая бифуркация (жесткое возбуждение автоколебаний). В этом случае при бифуркационном значении параметра устойчивый фокус теряет устойчивость из-за «влипания» в него неустойчивого предельного цикла (рис. 8.7). Фокус становится неустойчивым, а аттрактором при этом может стать предельный цикл большой амплитуды.
Рис. 8.7.
Докритическая (субкритическая) бифуркация Андронова – Хопфа.
Жесткое возбуждение автоколебаний. «Локальные события» при изменении
параметра при переходе через бифуркационное значение: устойчивый фокус и
неустойчивый предельный цикл пунктир, (а)
при уменьшении параметра с переходят в центр (б), а затем в неустойчивый фокус (в). Внешний большой устойчивый предельный цикл находится за
пределами локальной системы, в которой происходит бифуркация. Устойчивый фокус (неустойчивый предельный цикл) Центр Неустойчивый фокус (устойчивый предельный цикл)

«Модельной» системой (см. лекция 6), описывающей рождение предельного цикла при жестком возбуждении, является система:
![]()
![]()
Приравняв правую часть первого уравнения нулю, получим стационарные значения r:
![]()
![]()
Ветвь r = 0 устойчива при c < 0 и неустойчива при c > 0.
Рис 8.8. Фазовые траектории
в окрестности «черной дыры» при разных значениях параметра c. Устойчивый фокус Устойчивый фокус, неустойчивый предельный цикл, устойчивый предельный цикл Неустойчивый фокус, устойчивый предельный цикл
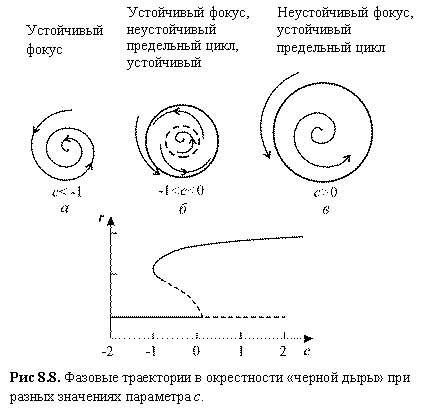
При с > –1
стационарное решение ![]()
При –1 < с < 0 стационарное решение ![]()
Рассмотрим, что произойдет, если
двигаться по параметру с, начиная с
отрицательных значений (Рис.8.8). Первоначально имеется единственное устойчивое
стационарное состояние r = 0,
колебаний нет. При c > –1
существует также устойчивый предельный цикл, но система не покидает своего
устойчивого стационарного состояния. Однако после того как с становится
положительным, стационарное состояние становится неустойчивым, и происходит
резкий скачок к устойчивому предельному циклу. В системе начинаются колебания
сразу большой амплитуды. Если двигаться от положительных значений с к
отрицательным, колебания большой амплитуды сохраняются до тех пор, пока с
не станет меньше –1, а затем внезапно исчезнут. Таким образом при –1 < с < 0 могут существовать два различных типа поведения.
Какой из них реализуется, зависит от предыстории системы. Такой феномен
называется эффектом гистерезиса.
При увеличении параметра с и его переходе через ноль скачком возникают устойчивые автоколебания конечной амплитуды и частоты. Для промежуточных значений параметра с существуют два типа устойчивого поведения (два аттрактора) - устойчивое стационарное состояние и устойчивый предельный цикл.
Винфри (Winfree A.T.) назвал области, в которых возможны два режима: устойчивая точка покоя и предельный цикл, - черной дырой (рис. 8.8 б). В этой области параметров можно так приложить возмущение к колебательной системе, что она попадет в область притяжения точки покоя, что приведет к прекращению колебаний. В частности, это показано для уравнений Ходжкина–Хаксли, моделирующих проведение нервного импульса (см. ниже).
Брюсселятор.
Простейшим классическим примером
существования автоколебаний в системе химических реакций является тримолекулярная
модель «Брюсселятор», предложенная в Брюсселе Пригожиным и Лефевром (1965).
Основной целью при изучении этой модели было установление качественных типов
поведения, совместимых с фундаментальными законами химической и биологической
кинетики.
В этом смысле блюсселятор играет роль базовой модели, такую же как гармонический осциллятор в физике, или модели Вольтерра в динамике популяций. Во 2-й части лекций мы остановимся на пространственно‑временных свойствах распределенной системы, локальным элементом которой является брюсселятор. Здесь мы рассмотрим свойства брюсселятора как автоколебательной системы.
Брюсселятор
содержит простейшую реализацию кубической нелинейности посредством химической реакции
2X + Y ® 3X (8.5)
Хотя тримолекулярная стадия в химической кинетике
не столь распространена, как бимолекулярные процессы, выражения для скорости
ряда биохимических реакций в определенных случаях можно свести к кубическому
виду. В качестве примера приведем следующую последовательность ферментативных
реакций:
X + E ® EX
EX + Y ® XY
EXY + X ® EX2Y
Здесь предполагается что фермент E
имеет по крайней мере три каталитических центра, способных одновременно
фиксировать две молекулы X и одну
молекулу Y. Если
образующиеся комплексы распадаются с достаточно большой скоростью, а ферменты
присутствуют в небольших количествах, легко показать, что всю
последовательность реакций можно свести к одной стадии, дающей нелинейный член
типа X 2Y в выражении для скорости
реакции.
Брюсселятор представляет собой следующую схему гипотетических химических
реакций:
A X 2X +
Y 3X B + X Y + C X R
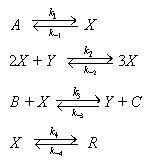
Здесь А, В — исходные вещества, C,
R — продукты, X, Y — промежуточные
вещества.
Пусть конечные продукты С и R немедленно удаляются из реакционного
пространства. Это означает, что обратные константы k-3 = k-4 = 0.
Если субстрат A находится в избытке, k-1 = 0.
Предположим также, что k-2 = 0.
Значения остальных констант положим равными единице. Тогда схема реакций 9.2 (в
случае точечной системы) описывается системой уравнений:
![]()
![]()
Модель
(8.5) имеет одну особую точку с координатами:
![]()
Исследуем стационарное решение (8.6) на устойчивость по
методу Ляпунова. Введем переменные, характеризующие отклонения от особой точки:
![]()
Линеаризованная система имеет вид:
![]()
![]()
Характеристическое уравнение
![]()
или
l2 + (A2 + 1 - B)l + A2 = 0
имеет корни:
![]()
Напомним, что особая точка является устойчивой, если действительные части корней характеристического уравнения отрицательны. Из выражения (8.7) видно, что при B < 1 + A2 особая точка (8.6) устойчива. Если же B > 1 + A2 особая точка становится неустойчивой, и у системы (8.5) появляется устойчивый предельный цикл. Значение B = 1 + A2 является бифуркационным. Если величина B лишь немного превосходит бифуркационный порог, автоколебания в системе носят квазигармонический характер. Таким образом, брюсселятор при выполнении условия
Рис. 8.9. Фазовый портрет
системы брюсселятор при B>1+A2
(а) и B<1+A2 (б).
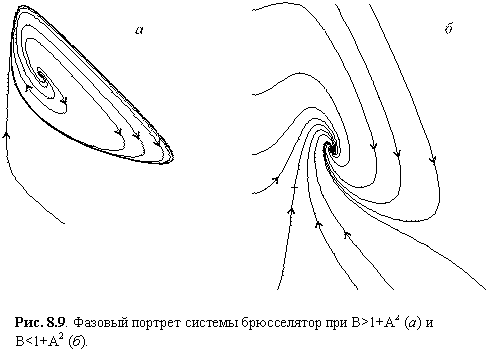
B > 1 + A2 (8.8)
является
автоколебательной системой. Фазовый портрет брюсселятора при разных значениях
параметров изображен на рис. 8.9
Здесь мы приведем краткий обзор нескольких «успешных» моделей колебательных биологических процессов. Более подробно некоторые колебательные процессы будут рассмотрены в лекциях 9, 11, 12.
Модель темновых процессов
фотосинтеза.
Одной из первых моделей, описывающих колебательный процесс в живой системе,
была модель темновых процессов фотосинтеза, предложенная и исследованная
Д.С.Чернавским с сотрудниками (1967). Модель является примером системы второго
порядка с квадратичными правыми частями, в которой возникают автоколебания
(существует предельный цикл) и допускает полное аналитической исследование
(Белюстина, 1967)
Рис. 8.10. Зависимость
поглощения кислорода и выделения углекислоты зеленым листом от времени. а
‑ при периодическом освещении; б - при непрерывном освещении

Известно,
что в условиях смены дня и ночи интенсивность фотосинтеза, то есть скорость
выделения кислорода и поглощения СО2 изменяется периодически
(рис.8.10 а). Если растение
поместить в условия непрерывной освещенности, то периодичность в интенсивности
фотосинтеза с периодом несколько часов сохраняется достаточно длительное время.
По-видимому, растение имеет свой внутренний ритм, синхронизованный с
периодическим внешним воздействием.
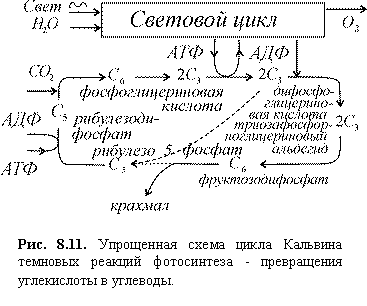
Напомним, что в процесс фотосинтеза входят световой и темновой циклы
химических реакций. Первый включает поглощение энергии квантов света и, через
ряд промежуточных стадий, приводит к образованию высокоэнергетических
восстановленных химических соединений и богатых энергией молекул АТФ. Эти вещества
употребляются в темновом цикле (цикле Кальвина), в котором свет непосредственно
не участвует. Здесь происходит восстановление углекислоты СО2 с
помощью веществ, богатых энергией, и доноров водорода, полученных в световом
цикле, и превращение ее в углеводы ‑ фруктозу и глюкозу (рис. 8.11).
В цикле участвуют углеводы с различным содержанием углерода (индекс внизу
означает число атомов углевода в молекуле). Все трехуглеродные сахара имеют
общее название триозы (с3), пятиуглеродные (с5) ‑ пентозы, шестиуглеродные
(с6) ‑ гексозы. Цикл замкнут, т.е. вещество, к
которому первоначально присоединяется углекислота (акцептор СО2,
обозначенный на рис. 8.8 символом с5) в результате реакции
регенерируется. Самые простые сахара ‑ триозы ‑ непосредственно
связаны со световым циклом, остальные сахара со световым циклом не связаны. Все
реакции в цикле, за исключением первичной фиксации СО2 на рибулезе,
бимолекулярные, и зависимость скорости реакции от концентрации описывается
членами второго порядка.
Рис. 8.11. Упрощенная схема
цикла Кальвина темновых реакций фотосинтеза ‑ превращения углекислоты
в углеводы.
Для
упрощения системы были выделены группы веществ, реакции между которыми
протекают быстро и обратимо, легкие сахара (трехуглеродистые углеводы) и более
тяжелые шестиуглеродные сахара. Суммарная концентрация первых обозначалась условно
с3, а вторых с6.
Предполагалось, что прибыль тяжелых сахаров с6 может
осуществляться за счет соединения двух легких с3. Их
убыль, так же как и убыль тяжелых сахаров, происходит в результате
бимолекулярного взаимодействия тяжелых и легких сахаров. Имеет место также
приток продукта с3 в сферу реакции за счет биохимически сходных процессов
(гликолиза, дыхания). Эти предположения приводят к системе уравнений:
Рис.
8.12. Фазовый портрет системы (8.9) при разных соотношениях
параметров (указаны в тексте). а – стационарное состояние (1,1) –
устойчивый фокус. б – (1,1) – неустойчивый фокус, жирная кривая –
предельный цикл
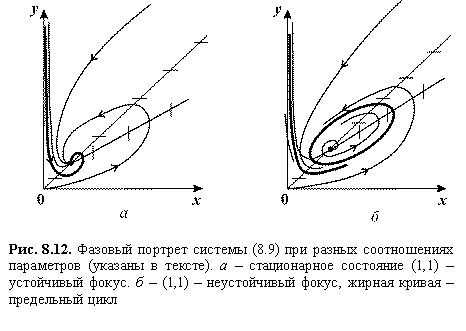
Переменные представляют собой нормированные концентрации легких (x) и тяжелых (y) сахаров. В положительном квадранте имеется одно состояние равновесия с координатами (1,1). Изоклины горизонтальных касательных определяются из уравнения
![]()
а изоклины вертикальных касательных – из уравнения
![]()
На рис. 8.12 изображены фазовые портреты системы. При ![]()
![]()
Колебания в гликолизе.
Классическим примером колебательной
биохимической реакции является гликолиз. В процессе гликолиза осуществляется
распад глюкозы и других сахаров, при этом соединения, содержащие шесть молекул
углерода, превращаются в трикарбоновые кислоты, включающие три молекулы
углерода. За счет избытка свободной энергии в процессе гликолиза на одну
молекулу шестиуглеродного сахара образуются две молекулы АТФ. Основную роль в
генерации наблюдаемых колебаний концентраций компонентов реакции:
фруктозо-6-фосфата, фруктозо-1,6-фосфата и восстановленного НАД (никотинаминадениндинуклеотид) играет
ключевой фермент гликолитического пути - фосфофруктокиназа (ФФК).
Рис. 8.13. Упрощенная
схема
реакций гликолиза
Активация
¯
[Гл] ® Ф6Ф® ФДФ
®
(x) (y)
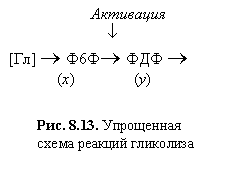 Упрощенная схема реакций
представлена на рис. 8.13:
Упрощенная схема реакций
представлена на рис. 8.13:
|
Рис. 8.13. Упрощенная схема
реакций гликолиза |
|
Активация ¯ [Гл] ® Ф6Ф® ФДФ
® (x) (y) |
На схеме [Гл] - глюкоза, Ф6Ф -
фруктозо-6-фосфат - субстрат ключевой реакции, ФДФ ‑
продукт этой реакции, который является субстратом в следующей стадии. Обе
реакции катализируются ферментами. В безразмерных переменных система описывающих
реакции уравнений может быть записана в виде:

Здесь зависимости скоростей реакций от переменных
записаны в форме Михаэлиса-Ментен (Моно), как это было представлено в уравнении
(8.10).Если выполняются условия: Kmx>>x, Kmy>>y,
можно произвести замену переменных
![]()
Опустив штрихи у новых переменных, получим систему в
безразмерном виде :
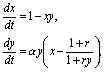
где
![]()
Кинетика изменений переменных и фазовые портреты системы
(8.10) при разных значениях параметров представлены на рис. 8.14
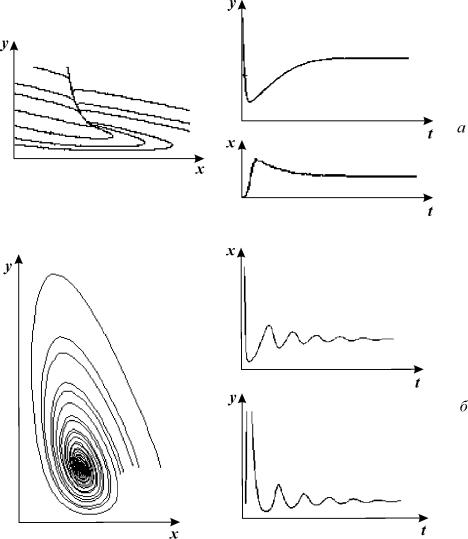
Рис. 8.14. Модель гликолиза (8.10). Кинетика изменений
концентраций фруктозо‑6‑фосфата (х) и фруктозодифосфата (у)
(справа) и фазовый портрет системы (слева) при разных значениях параметров
системы, а ‑ бесколебательный процесс (узел на фазовой плоскости),
α = 0.25; r = 1.
б – затухающие колебания (устойчивый фокус на фазовой плоскости)
α = 4; r = 0.2
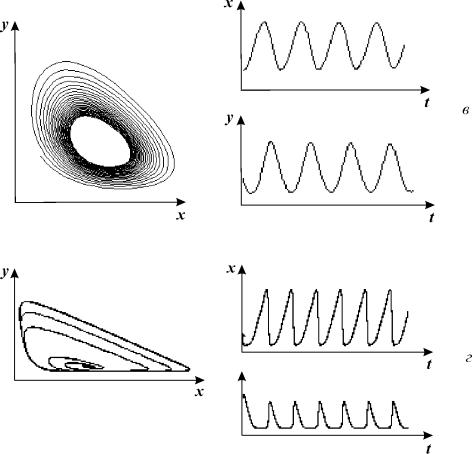
Рис. 8.14. Модель гликолиза (8.10). Кинетика изменений
концентраций фруктозо‑6‑фосфата (х) и фруктозодифосфата (у)
(справа) и фазовый портрет системы (слева) при разных значениях параметров
системы, в – колебания с постоянной амплитудой и фазой,
близкие к гармоническим (предельный цикл на фазовой плоскости),
α = 6; r = 0.2.
г – релаксационные колебания с постоянной амплитудой и фазой,
(предельный цикл почти треугольной формы на фазовой плоскости )
α = 8; r = 0.5
Интересно,
что колебательные реакции в системе гликолиза были сначала предсказаны на
математической модели (Higgins, 1964), и лишь после этого зарегистрированы
экспериментально с помощью метода дифференциальной спектрофотометрии в
лаборатории Б. Чанса
(1966).
Внутриклеточные колебания кальция.
Во многих типах живых клеток наблюдаются колебания внутриклеточной
концентрации кальция, период которых может варьировать от 0,5 до 10 мин.
Простейшая схема процессов, приводящих к гармонально обусловленным колебаниям
кальция, основой которых служит кальций индуцированный выходакальция из клетки,
приведена на рис. 8.15. Такие колебания впервые наблюдались Эндо с соавторами
(1970) на клетках скелетных мышц, Фабиато (1975) на клетках
саркоплазматического ретикулума сердца быка, и позднее — многими другими
исследователями.
Схема и модель процессов, предложена и описана в
[Dupont and Goldbetter (1989, 1994)]. Рассматриваются приток и отток кальция в
клетку через плазматическую мембрану (константы скоростей v1 и v2,
соответственно); гормонально активируемое освобождение кальция из пула
(скорость v3); активный транспорт цитозольного
кальция в пул, (v4), освобождение кальция из
пула, активируемое цитозольным кальцием (v5); свободный отток кальция из
пула в цитозоль (v6). Модель состоит из двух
дифференциальных уравнений
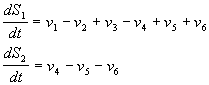
Рис.
8.15: Схема процессов, приводящих к внутриклеточным
колебаниям кальция (Dupont, Goldbeter, 1983). IP3 ‑
рецептор, стимулирующий колебания
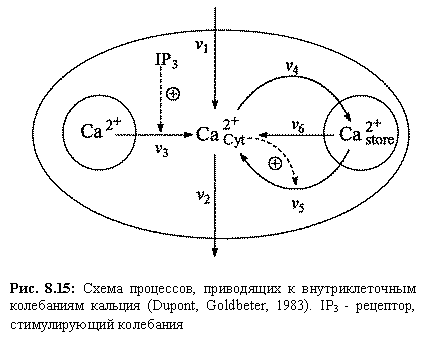
Здесь
S1 ‑ концентрация кальция в цитозоле, S1‑ концентрация
кальция в гормонально чувствительном пуле.
Выражения для величин скоростей были предложены в Simogyi, Stuckin (1991):
![]()
Модель предсказывает колебания концентрации кальция во времени, по форме близкие к экспериментальным (рис. 8.16).
Рис. 8.16: Модель
внутриклеточных колебаний кальция. Кинетика концентрации Ca при
разных значениях параметров. a ‑ a b
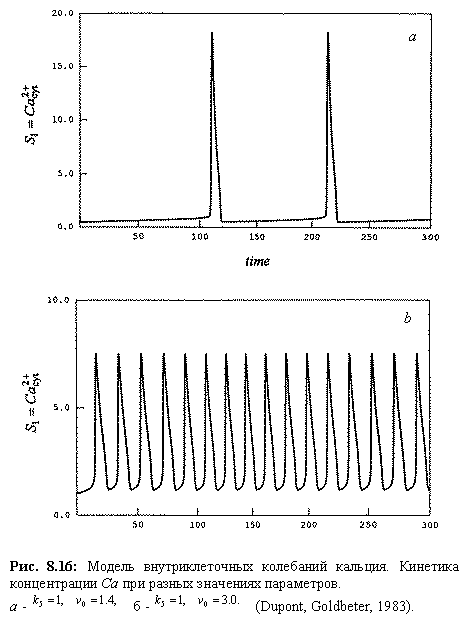
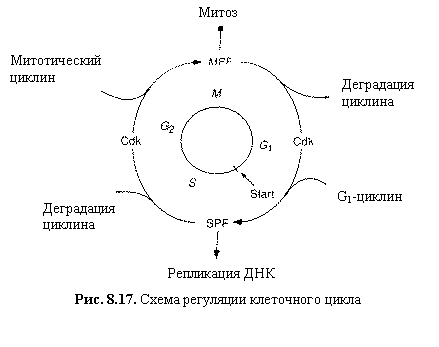
Рис. 8.17. Схема
регуляции клеточного цикла Репликация ДНК Митоз Деградация циклина Деградация циклина G1-циклин Митотический циклин
Клеточные циклы.
В
процессе жизненного цикла клетка удваивает свое содержимое и делится на две. В
организме млекопитающего для поддержания жизни производятся ежесекундно
миллионы новых клеток. Нарушение регуляции пролиферации клеток проявляется как
онкологическое заболевание. Этим вызван большой интерес к изучению и
моделированию механизмов регуляции клеточного деления.
Схема клеточного цикла изображена на рис. 8.17. Клеточный цикл состоит из
двух периодов: митоз (М‑фаза)
включает разделение предварительно удвоенного ядерного материала, деление ядра
и деление самой клетки ‑ цитокинез и занимает около часа. Значительно
более длительный период между двумя митозами занимает интерфаза, включающая
стадию роста G1, фазу
репликации ДНК (S), фазу подготовки к
делению G2. Клеточный цикл
(рис.8.17) регулируется генами и белками-ферментами двух основных классов.
Рис. 8.18. Схема стадий
клеточного цикла
Циклин-зависимые протеин‑киназы (Cdk) индуцируют последовательность процессов путем фосфорилирования отдельных белков. Циклины, которые синтезируются и деградируют при каждом новом цикле деления, связываются с молекулами Cdk и контролируют их способность к фосфорилированию, без циклина Cdk не активны. Количество этих молекул‑регуляторов различно в разного вида клетках.
В делении дрожжевой клетки основные роли играют один Cdk и девять циклинов, которые образуют девять разных циклин‑Cdk комплексов. У гораздо более сложно организованных млекопитающих изучено шесть Cdk и полтора десятка циклинов. Контроль выхода клетки из G1, и G2 фаз осуществляют промотор‑фактор S‑фазы (SPF) и промотор‑фактор M‑фазы (MPF), представляющие собой гетеродимеры. Cуществует особая контрольная точка клеточного цикла (Start), с которой заканчивается рост (G1 фаза) и начинается процесс синтеза ДНК.
Простая модель процесса предложена Тайсоном (Tyson, 1995). Постулируется существование фактора транскрипции SBF, который может быть в активной Sa и пассивной Si форме. Он переходит в активную форму под действием циклина Cln (N) и Start‑киназы (Cdc28-Cln3) (A) и инактивируется другим веществом (Е). Циклин продуцируется путем активации SBF и деградирует. SBF активируется Chu и Start-киназой и инактивируется фосфатазой. Безразмерная модель процессов имеет вид:
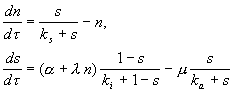
Модель имеет одно или три стационарных решения (два устойчивых) в
зависимости от значений параметров, и при увеличении параметра a (в
процессе роста клетки) описывает переключение системы из G1 в S фазу.
Добавление двух уравнений сходного вида позволяет описать также
переключение из G2 в фазу
митоза М. Полная модель, учитывающая
и другие регуляторные ферменты в фосфорилированной и дефосфорилированной форме
содержит 9 нелинейных уравнений (Novak, Tyson 1993) и хорошо описывает кинетику
деления ооцитов Xenopus. При соответствующем
подборе параметров она применима к описанию деления других типов клеток.
Большое количество работ было посвящено попыткам моделирования
периодического воздействия на клеточный цикл с целью оптимизации параметров
рентгено‑ радио‑ или хемотерапии при воздействии на клетки
онкологических опухолей.
В современной литературе по математической биологии рассмотрены сотни автоколебательных систем на разных уровнях организации живой природы. Несомненно, колебательный характер процессов — эволюционное изобретение природы, и их функциональная роль имеет несколько разных аспектов.
Во-первых колебания позволяют разделить процессы во времени, когда в одном компартменте клетки протекает сразу несколько различных реакций, организуя периоды высокой и низкой активности отдельных метаболитов. Во-вторых, характеристики колебаний, их амплитуда и фаза, несут определенную информацию и могут играть регуляторную роль в каскадах процессов, проходящих на уровне клетки и живого организма. Наконец, колебательные (потенциально или реально) системы служат локальными элементами распределенных активных сред, способных к пространственно‑временной самоорганизации, в том числе к процессам морфогенеза. Эту роль колебательных систем мы подробно рассмотрим во второй части лекций.
Литература
Андронов А.А., Витт А.А., Хайкин С.Э. Теория
колебаний. М., Наука, 1981.
Белюстина Л.Н., Кокина Г.А. Качественное исследовани
е уравнений фотосинтеза. – В сб. Колебательные процессы в биологических и
химических системах. М., Наука, 1967
Гленсдорф П., Пригожин И. Термодинамическая
теория структуры, стабильности и флуктуаций. М., 1978
Николис Ж, Пригожин И. Самоорганизация в
неравновесных системах. М., Мир., 1979
Чернавский Д.С., Чернавская Н.М. О колебаниях
в темновых реакциях фотосинтеза. – В сб. Колебательные процессы в биологических
и химических системах. М., Наука, 1967
Dupont
G. and Goldbetter A. Theoretical insights into the origin of
signal-induced calcium oscillations, in Goldbeter A. (ed). Cell to Cell
signalling: From experiments to theoretical models. Acad. Press, London,
pp. 461-474, 1989
Dupont
G. and Goldbetter A. Oscillations and waves of citosolic calsium: insights
from theoretical models. Bioessays, 14, 485-493, 1992
Higgins
J.A. A chemical mechanism for oscillations in glicolitic intermediates in yeast
cells. Proc. Nat. Acad. Sci. USA, v.51, 1954
Higgins
J.A. The theory of oscillating reactions. Ing. Chem. V.59, N5, 1967
Hopf
E. Abzweilung einer periodischen Losung von einer stationaren Losung eines
Differentialsystems (bifurcation of a periodic solusion from stationary
solution of a system of differential equations) Ber. Math-phys. Kl. Sachs.
Akad. Wiss. Leipzig 94, 3-22, 1942
Novak B, Tyson
J.J. Modeling the cell division cycle: M-phase trigger, oscillation and size
control, J. Theor. Biol. 165, 101-104, 1993
Somogyi R,
Stuckin J.W. Hormone-induced calcium oscillations in liver cells can be
explained by a simple one pool model.
J. Biol. Chem.266, 11068-11077, 1991
Prigozhine I.R., Lefebre R. Simmetry
breaking instabilities indissipative systems.J. Che. Phys., 48, 1665-1700, 1968
Winfree A.T. The Geometry of Biological Time. Springer, 1980
