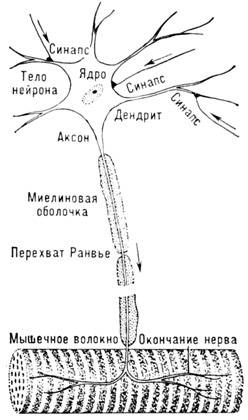
Нервный импульс - волна возбуждения, к-рая
распространяется по нервному волокну и служит для передачи информации от периферич.
рецепторных (чувствительных) окончаний к нервным центрам, внутри центр. нервной
системы и от неё к исполнительным аппаратам - мышцам и железам. Прохождение
Н. и. сопровождается переходными электрич. процессами, к-рые можно зарегистрировать
как внеклеточными, так и внутриклеточными электродами.
Генерацию, передачу и переработку Н. и. осуществляет
нервная система. Осн. структурным элементом нервной системы высших организмов
является нервная клетка, или нейрон, состоящий из тела клетки и многочисл. отростков
- дендритов (рис. 1). Один из отростков у нериферич. нейронов имеет большую
длину - это нервное волокно, или аксон, протяжённость к-рого ~ 1 м, а толщина
от 0,5 до 30 мкм. Различают два класса нервных волокон: мякотные (мие-линизированные)
и безмякотные. У мякотных волокон имеется миелиновая оболочка, образованная
спец. мембраной, к-рая подобно изоляции накручивается на аксон. Протяжённость
участков сплошной миелиновой оболочки составляет от 200 мкм до 1 мм, они прерываются
т. н. перехватами Ранвье шириной 1мкм. Миелиновая оболочка играет роль изоляции;
нервное волокно на этих участках пассивно, электрически активна только мембрана
в перехватах Ранвье. Безмякотные волокна не имеют изолир. участков; их структура
однородна по всей длине, а мембрана обладает электрич. активностью по всей поверхности.
Нервные волокна заканчиваются на телах или ден-дритах
др. нервных клеток, но отделены от них проме-
жутком шириной ~ 10 нм. Эта область контакта
двух клеток наз. синапсом. Входящая в синапс мембрана аксона наз. пресинаптической,
а соответствующая мембрана дендритов или мышцы - пост-синаптической (см. Клеточные
структуры).
В нормальных условиях по нервному волокну постоянно
бегут серии Н. и., возникающих на дендритах или теле клетки и распространяющихся
по аксону в направлении от тела клетки (аксон может проводить Н. и. в обоих
направлениях). Частота этих периодич. разрядов несёт информацию о силе вызвавшего
их раздражения; напр., при умеренной активности частота ~ 50-100 импульсов/с.
Существуют клетки, к-рые разряжаются с частотой ~ 1500 импульсов/с.
Скорость распространения Н. и. u
зависит от типа нервного волокна и его диаметра d, u
~ d1/2. В тонких волокнах нервной системы человека u
~ 1 м/с, а в толстых волокнах u
~ 100-120 м/с.
Каждый Н. и. возникает в результате раздражения
тела нервной клетки или нервного волокна. Н. и. всегда имеет одни и те же характеристики
(форму и скорость) независимо от силы раздражения, т. е. при подпороговом раздражении
Н. и. не возникает совсем, а при надпороговым - имеет полную амплитуду.
После возбуждения наступает рефракторный период,
в течение к-рого возбудимость нервного волокна снижена. Различают абс. рефракторный
период, когда волокно нельзя возбудить никакими раздражителями, и относит. рефракторный
период, когда возбуждение возможно, но его порог оказывается выше нормы. Абс.
рефракторный период ограничивает сверху частоту передачи Н. и. Нервное волокно
обладает свойством аккомодации, т. е. привыкает к постоянно действующему раздражению,
что выражается в постепенном повышении порога возбудимости. Это приводит к снижению
частоты Н. и. и даже к их полному исчезновению. Если сила раздражения нарастает
медленно, то возбуждения может не произойти даже после достижения порога.
Рис.1. Схема строения нервной клетки.
Вдоль нервного волокна Н. и. распространяется
в виде волны электрич. потенциала. В синапсе происходит смена механизма распространения.
Когда Н. и. достигает пресинаптич. окончания, в синаптич. щель выделяется активное
хим. вещество - м е д и а т о р. Медиатор диффундирует через синаптич. щель
и меняет проницаемость постсинаптич. мембраны, в результате чего на ней возникает
потенциал, вновь генерирующий распространяющийся импульс. Так действует хим.
синапс. Встречается также электрич. синапс, когда след. нейрон возбуждается
электрически.
Возбуждение Н. и. Физ. представления о
появлении электрич. потенциалов в клетках основаны на т. н. мембранной теории.
Клеточные мембраны разделяют растворы электролита разной концентрации и обладают
из-бират. проницаемостью для нек-рых ионов. Так, мембрана аксона представляет
собой тонкий слой липидов и белков толщиной ~ 7 нм. Её электрич. сопротивление
в состоянии покоя ~ 0,1 Ом.м2, а ёмкость ~ 10 мф/м2.
Внутри аксона высока концентрация ионов К+ и мала концентрация ионов
Na+ и Сl-, а в окружающей среде - наоборот.
В состоянии покоя мембрана аксона проницаема
для ионов К+. Из-за разницы концентраций C0K
во внеш. и С во внутр. растворах на мембране устанавливается калиевый мембранный
потенциал
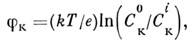
где Т - абс. темп-pa, е - заряд
электрона. На мембране аксона действительно наблюдается потенциал покоя ~ -60
мВ, соответствующий указанной ф-ле.
Ионы Na+ и Сl- проникают
через мембрану. Для поддержания необходимого неравновесного распределения ионов
клетка использует систему активного транспорта, на работу к-рой расходуется
клеточная энергия. Поэтому состояние покоя нервного волокна не является термодинамически
равновесным. Оно стационарно благодаря действию ионных насосов, причём мембранный
потенциал в условиях разомкнутой цепи определяется из равенства нулю полного
электрич. тока.
Процесс нервного возбуждения развивается следующим
образом (см. также Биофизика ).Если пропустить через аксон слабый импульс
тока, приводящий к деполяризации мембраны,
то после снятия внеш. воздействия потенциал монотонно возвращается к исходному
уровню. В этих условиях аксон ведёт себя как пассивная электрич. цепь, состоящая
из конденсатора и пост. сопротивления.
Рис. 2. Развитие потенциала действия в нервном
волокне: а - подпороговое
(1) и надпороговое (2) раздражения;
б -мембранный отклик; при над-пороговом раздражении проявляется полный
потенциал действия; в - ионный
ток, протекающий через мембрану при
возбуждении; г - аппроксимация ионного
тока в простой аналитической модели.
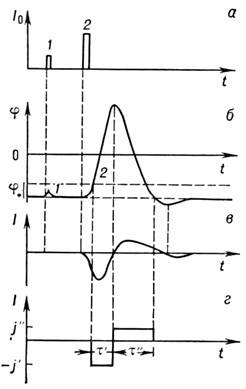
Если импульс тока превышает нек-рую пороговую
величину, потенциал продолжает изменяться и после выключения возмущения; потенциал
становится положительным и только потом возвращается к уровню покоя, причём
вначале даже несколько проскакивает его (область гиперполяризации, рис. 2).
Отклик мембраны при этом не зависит от возмущения; этот импульс наз. потенциалом
действия. Одновременно через мембрану течёт ионный ток, направленный сначала
внутрь, а потом наружу (рис. 2, в).
Феноменологич. истолкование механизма возникновения
Н. и. было дано А. Л. Ходжкином (A. L. Hodg-kin) и А. Ф. Хаксли (A. F. Huxley)
в 1952. Полный ионный ток слагается из трёх составляющих: калиевого, натриевого
и тока утечки. Когда потенциал мембраны сдвигается на пороговую величину j*
(~ 20мВ), мембрана становится проницаемой для ионов Na+. Ионы Na+
устремляются внутрь волокна, сдвигая мембранный потенциал, пока он не достигнет
величины равновесного натриевого потенциала:
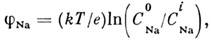
составляющего ~ 60 мВ. Поэтому полная амплитуда
потенциала действия достигает ~ 120 мВ. К моменту достижения макс. потенциала
в мембране начинает развиваться калиевая (и одновременно уменьшаться натриевая)
проводимость. В результате натриевый ток сменяется
на калиевый, направленный наружу. Этот ток соответствует уменьшению потенциала
действия.
Установлены эмпирич. ур-ния для описания натриевого
и калиевого токов. Поведение мембранного потенциала при пространственно однородном
возбуждении волокна определяется ур-нием:

где С - ёмкость мембраны, I - ионный
ток, слагающийся из калиевого, натриевого и тока утечки. Эти токи определяются
пост. эдс jK,
jNa
и jl
и проводимостями gK, gNa и gl:
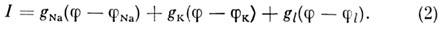
Величину gl считают постоянной,
проводимости gNa и gK описывают с
помощью параметров m, h и п:
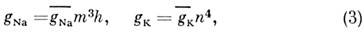
gNa, gK -
постоянные; параметры т, h и п удовлетворяют линейным ур-ниям
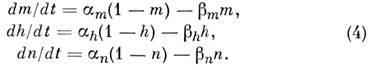
Зависимость коэф. a
и b от мембранного
потенциала j
(рис. 3) выбирают из условия наилучшего совпадения
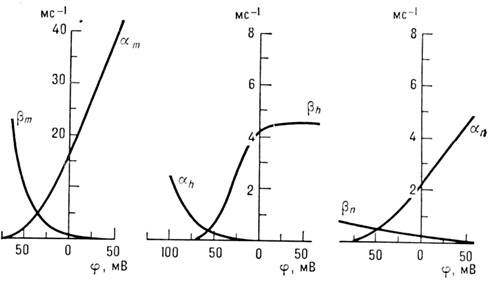
Рис. 3. Зависимость коэффициентов a
и b
от мембранного потенциала.
расчётных и измеряемых кривых I(t).
Этими же соображениями вызван выбор параметров. Зависимость стационарных значений
т, h и п от мембранного потенциала приведена на рис. 4. Существуют
модели с большим числом параметров. Т. о., мембрана нервного волокна представляет
собой нелинейный ионный проводник, свойства к-рого существенно зависят от электрич.
поля. Механизм генерации возбуждения изучен плохо. Ур-ния Ходжкина -Хаксли дают
лишь удачное эмпирич. описание явления, за к-рым нет конкретной физ. модели.
Поэтому важной задачей является изучение механизмов протекания электрич. тока
через мембраны, в частности через управляемые элект- рич. полем ионные каналы.
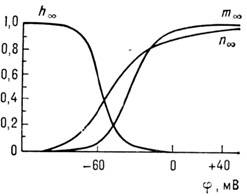
Рис. 4. Зависимость стационарных значений
т, h и п от мембранного
потенциала.
Распространение Н. и. Н. и. может распространяться
вдоль волокна без затухания и с пост. скоростью. Это связано с тем, что необходимая
для передачи сигнала энергия не поступает из единого центра, а черпается на
месте, в каждой точке волокна. В соответствии с двумя типами волокон существуют
два способа передачи Н. и.: непрерывный и сальтаторный (скачкообразный), когда
импульс движется от одного перехвата Ранвье к другому,
перепрыгивая через области миелиновой изоляции.
В случае немиелинизир. волокна распределение
мембранного потенциала j(x,
t)определяется ур-нием:
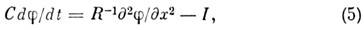
где С - ёмкость мембраны, приходящаяся
на единицу длины волокна, R - сумма продольных (внутриклеточного и внеклеточного)
сопротивлений на единицу длины волокна, I - ионный ток, протекающий через
мембрану волокна единичной длины. Электрич. ток I является функционалом
от потенциала j,
к-рый зависит от времени t и координаты х. Эта зависимость определяется
ур-ниями (2) -(4).
Вид функционала I специфичен для биологически
возбудимой среды. Однако ур-ние (5), если отвлечься от вида I, имеет
более общий характер и описывает многие физ. явления, напр. процесс горения.
Поэтому передачу Н. и. уподобляют горению порохового шнура. Если в бегущем пламени
процесс поджигания осуществляется за счёт теплопроводности, то в Н. и. возбуждение
происходит при помощи т. н. локальных токов (рис. 5).
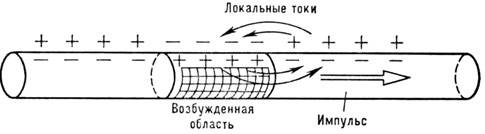
Рис. 5. Локальные токи, обеспечивающие распространение
нервного импульса.
Ур-ния Ходжкина - Хаксли для распространения
Н. и. решались численно. Полученные решения вместе с накопленными эксперим.
данными показали, что распространение Н. и. не зависит от деталей процесса возбуждения.
Качеств. картину распространения Н. и. можно получить при помощи простых моделей,
отражающих лишь общие свойства возбуждения. Такой подход позволил рассчитывать
скорость и форму Н. и. в однородном волокне, их изменение при наличии неоднород-ностей
и даже сложные режимы распространения возбуждения в активных средах, напр. в
сердечной мышце. Существует неск. матем. моделей подобного рода. Простейшая
из них такова. Ионный ток, протекающий через мембрану при прохождении Н. и.,
является знакопеременным: вначале он течёт внутрь волокна, а потом наружу. Поэтому
его можно аппроксимировать кусочно-постоянной функцией (рис. 2, г). Возбуждение
происходит, когда мембранный потенциал сдвигается на пороговую величину j*.
В этот момент возникает ток, направленный внутрь волокна и равный по модулю
j'. Спустя время t'
ток меняется на противоположный, равный j''. Эта фаза продолжается
в течение времени ~ t''.
Автомодельное
решение ур-ния (5) можно найти как функцию переменной t = х/u, где u
- скорость распространения Н. и. (рис. 2, б).
В реальных волокнах время t'
достаточно велико, поэтому только оно определяет скорость u, для к-рой справедлива ф-ла: 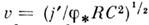 . Учитывая, что j' ~ ~d, R ~ d2 и С ~
d, где d - диаметр волокна, находим в согласии с экспериментом,
что u ~ d1/2. С помощью кусочно-постоянной аппроксимации находят форму потенциала
действия.
. Учитывая, что j' ~ ~d, R ~ d2 и С ~
d, где d - диаметр волокна, находим в согласии с экспериментом,
что u ~ d1/2. С помощью кусочно-постоянной аппроксимации находят форму потенциала
действия.
Ур-ние (5) для распространяющегося Н. и. в действительности
допускает два решения. Второе решение оказывается неустойчивым; оно даёт Н.
и. со значительно меньшей скоростью и амплитудой потенциала. Наличие второго,
неустойчивого, решения имеет аналогию в теории горения. При распространении
пламени с боковым теплоотводом также возможно возникновение неустойчивого режима.
Простую аналитич. модель Н. и. можно усовершенствовать, учитывая дополнит. детали.
При изменении сечения и при ветвлении нервных
волокон прохождение Н. и. может быть затруднено или даже полностью блокировано.
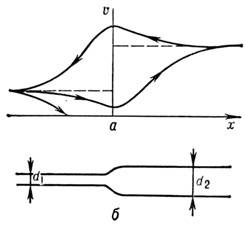
В расширяющемся волокне (рис. 6) скорость импульса по мере приближения к расширению
убывает, а после расширения начинает расти, пока не выйдет на новое стационарное
значение. Замедление Н. и. тем сильнее, чем больше разница в сечениях. При достаточно
большом расширении Н. и. останавливается. Существует критич. расширение волокна,
к-рое задерживает Н. и.
При обратном движении Н. и. (из широкого волокна
в узкое) блокирования не происходит, но изменение скорости носит противоположный
характер. При подходе к сужению скорость Н. и. увеличивается, а затем начинает
спадать до нового стационарного значения. На графике скорости (рис., 6а)
получается своего рода петля гистерезиса.
Рие. 6. Прохождение нервных импульсов по расширяющемуся
волокну: а - изменение скорости импульса в зависимости
от его направления; б -схематическое изображение
расширяющегося волокна.
Другой тип неоднородности - ветвление волокон.
В узле ветвления возможны разл. варианты прохождения и блокирования импульсов.
При несинхронном подходе Н. и. условие блокирования
зависит от временного сдвига. Если временной сдвиг между импульсами мал, то
они помогают друг другу проникнуть в широкое третье волокно. Если сдвиг достаточно
велик, то нервные импульсы мешают друг другу. Связано это с тем, что Н. и., подошедший
первым, но не сумевший возбудить третье волокно, частично переводит узел в рефракторное
состояние. Кроме того, возникает эффект синхронизации: по мере приближения Н.
и. к узлу их запаздывание друг относительно друга уменьшается.
Взаимодействие Н. и. Нервные волокна в
организме объединены в пучки или нервные стволы, образующие подобие многожильного
кабеля. Все волокна в пучке представляют собой самостоят. линии связи, но имеют
один общий "провод" - межклеточную жидкость. Когда по любому из
волокон бежит нервный импульс, он создаёт в межклеточной жидкости электрич. поле, к-рое
влияет на мембранный потенциал соседних волокон. Обычно такое влияние пренебрежимо
мало и линии связи работают без взаимных помех, но оно проявляется в пато-логич.
и искусств. условиях. Обрабатывая нервные стволы спец. хим. веществами, удаётся
наблюдать не только взаимные помехи, но и передачу возбуждения в соседние волокна.
Известны эксперименты по взаимодействию двух
нервных волокон, помещённых в ограниченный объём внеш. раствора. Если по одному
из волокон бежит нервный импульс, то одновременно изменяется возбудимость второго волокна.
Изменение проходит три стадии. Вначале возбудимость второго волокна падает (повышается
порог возбуждения). Это уменьшение возбудимости опережает потенциал действия,
бегущий по первому волокну, и длится примерно до тех пор, пока потенциал в первом
волокне не достигнет максимума. Затем возбудимость растёт, эта стадия совпадает
по времени с процессом уменьшения потенциала в первом волокне. Возбудимость
ещё раз уменьшается, когда в первом волокне происходит небольшая гиперполяризация
мембраны.
При одноврем. прохождении нервных импульсов по двум волокнам
иногда удавалось достигнуть их синхронизации. Несмотря на то что собств. скорости
нервных импульсов в разных волокнах различны, при их
одноврем. возбуждении мог возникнуть коллективный Н. и. Если собств. скорости
были одинаковы, то коллективный импульс имел меньшую скорость. При заметном
отличии собств. скоростей коллективная скорость имела промежуточное значение.
Синхронизоваться могли лишь нервные импульсы, скорости к-рых отличались не слишком сильно.
Матем. описание этого явления даётся системой
ур-ний для мембранных потенциалов двух параллельных волокон j1
и j2:
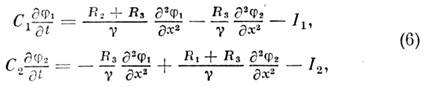
где R1 и R2
- продольные сопротивления первого и второго волокон, R3
- продольное сопротивление внешней среды, g
= R1R2 + R1R3
+ R2R3. Ионные токи I1
и I2 можно описать той или иной моделью нервного возбуждения.
При использовании простой аналитич. модели решение
приводит к след. картине. Когда возбуждается одно волокно, в соседнем наводится
знакопеременный мембранный потенциал: вначале волокно гиперполяри-зуется, затем
деполяризуется и, наконец, ещё раз ги-перполяризуется. Эти три фазы соответствуют
понижению, повышению и новому понижению возбудимости во-локна. При нормальных
значениях параметров сдвиг мембранного потенциала во второй фазе в сторону деполяризации
не достигает порога, поэтому передачи возбуждения в соседнее волокно не происходит.
При одноврем. возбуждении двух волокон система (6) допускает совместное автомодельное
решение, которое соответствует двум нервным импульсам, движущимся с одинаковой скоростью на
пост. расстоянии друг от друга. Если впереди находится медленный Н. и., то он
притормаживает быстрый импульс, не выпуская его вперёд; оба движутся с относительно
малой скоростью. Если же впереди находится быстрый II. и., то он подтягивает
за собой медленный импульс. Коллективная скорость оказывается близкой к собств.
скорости быстрого импульса. В сложных нейронных структурах возможно появление
автоволи.
Возбудимые среды. Нервные клетки в организме
объединены в нейронные сети, к-рые в зависимости от частоты ветвления волокон
разделяют на редкие и густые. В редкой сети отд. элементы возбуждаются независимо
друг от друга и взаимодействуют только в узлах ветвления, как описано выше.
В густой сети возбуждение охватывает сразу много
элементов, так что их детальная структура и способ соединения между собой оказываются
несущественными. Сеть ведёт себя как непрерывная возбудимая среда, параметры
к-рой определяют возникновение и распространение возбуждения.
Возбудимая среда может быть трёхмерной, хотя
чаще её рассматривают как двумерную поверхность. Возбуждение, возникшее в к--л.
точке поверхности, распространяется во все стороны в виде кольцевой волны. Волна
возбуждения может огибать препятствия, но не может от них отражаться, не отражается
она и от границы среды. При столкновении волн между собой происходит их взаимное
уничтожение; пройти друг сквозь друга эти волны не могут из-за наличия позади
фронта возбуждения рефракторной области.
Примером возбудимой среды является сердечный
нервно-мышечный синцитий - объединение нервных и мышечных волокон в единую проводящую
систему, способную передавать возбуждение в любом направлении. Нервно-мышечные
синцитии сокращаются синхронно, подчиняясь волне возбуждения, к-рую посылает
единый управляющий центр - водитель ритма. Единый ритм иногда нарушается, возникают
аритмии. Один из таких режимов наз. трепетанием предсердий: это автономные сокращения,
вызванные циркуляцией возбуждения вокруг препятствия, напр.
верхней или нижней вены. Для возникновения подобного режима периметр препятствия
должен превышать длину волны возбуждения, равную в предсердии человека ~ 5 см.
При трепетании происходит пе-риодич. сокращение предсердий с частотой 3-5 Гц.
Более сложный режим возбуждения представляет собой фибрилляция желудочков сердца,
когда отд. элементы сердечной мышцы начинают сокращаться без внеш. команды и
без связи с соседними элементами с частотой ~ 10 Гц. Фибрилляция приводит к
прекращению циркуляции крови.
Возникновение и поддержание спонтанной активности
возбудимой среды неразрывно связаны с возникновением источников волн. Простейший
источник волн (группа спонтанно возбуждающихся клеток) может обеспечить периодич.
пульсацию активности, так устроен водитель ритма сердца.
Источники возбуждения могут возникать и за счёт
сложной пространств. организации режима возбуждения, напр. ревербератор типа
вращающейся спиральной волны, появляющийся в простейшей возбудимой среде. Другой
вид ревербератора возникает в среде, состоящей из элементов двух типов с разными
порогами возбуждения; ревербератор периодически возбуждает то одни, то другие
элементы, меняя при этом направление своего движения и порождая плоские волны.
Третий вид источника - ведущий центр (источник эха), к-рый появляется в среде, неоднородной по реф-ракторности или порогу возбуждения. В этом случае на неоднородности возникает отражённая волна (эхо). Наличие подобных источников волн приводит к появлению сложных режимов возбуждения, исследуемых в теории автоволн.

В. С. Маркин
|
|
